Offering multiple perspectives from many fields of human inquiry that may move all of us toward a more integrated understanding of who we are as conscious beings.
Ed Yong - The Unique Merger That Made You (and Ewe, and Yew): One Freakish Event Responsible for All Sophisticated Life?
From Nautilus, this is a very cool article by Ed Yong (blogger at the National Geographic's Phenomena: Not Exactly Rocket Science blog) on the emergence of eukaryotic cells, an enormous leap in complexity and arguably one of the "most important event in the history of life on Earth."
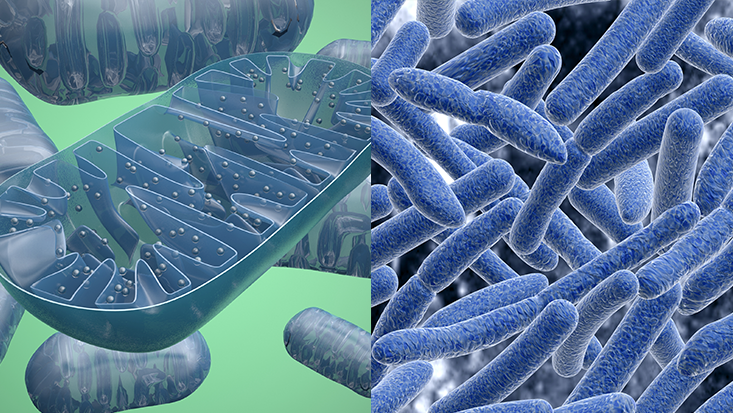
The emergence of eukaryotic life also brought along with it the emergence of mitochondria, the cell's "power plant" in that it generates most of the adenosine triphosphate (ATP) used for energy.
Interestingly, unlike other evolutionary innovations, the eukaryotic cell and mitochondria only appear once in the evolutionary timeline. Fortunately for us, they stuck around.
All sophisticated life on the planet Earth may owe its existence to one freakish event.
By Ed Yong | September 4, 2014
ILLUSTRATION BY GRACIA LAM
AT FIRST GLANCE, a tree could not be more different from the caterpillars that eat its leaves, the mushrooms sprouting from its bark, the grass growing by its trunk, or the humans canoodling under its shade. Appearances, however, can be deceiving. Zoom in closely, and you will see that these organisms are all surprisingly similar at a microscopic level. Specifically, they all consist of cells that share the same basic architecture. These cells contain a central nucleus—a command center that is stuffed with DNA and walled off by a membrane. Surrounding it are many smaller compartments that act like tiny organs, carrying out specialized tasks like storing molecules or making proteins. Among these are the mitochondria—bean-shaped power plants that provide the cells with energy. This combination of features is shared by almost every cell in every animal, plant, fungus, and alga, a group of organisms known as “eukaryotes.” Bacteria showcase a second, simpler way of building a cell—one that preceded the complex eukaryotes by at least a billion years. These “prokaryotes” always consist of a single cell, which is smaller than a typical eukaryotic one and bereft of internal compartments like mitochondria and a nucleus. Even though limited to a relatively simple cell, bacteria are impressive survival machines. They colonize every possible habitat, from miles-high clouds to the deep ocean. They have a dazzling array of biological tricks that allow them to cause diseases, eat crude oil, conduct electric currents, draw power from the Sun, and communicate with each other. Still, without the eukaryotic architecture, bacteria are forever constrained in size and complexity. Sure, they have their amazing skill sets, but it’s the eukaryotes that cover the Earth in forest and grassland, that navigate the planet looking for food and mates, that build rockets to Mars. The transition from the classic prokaryotic model to the deluxe eukaryotic one is arguably the most important event in the history of life on Earth. And in more than 3 billion years of existence, it happened exactly once. Life is full of complex structures that evolve time and again. Individual cells have united to form many-celled creatures like animals and plants on dozens of separate occasions. The same is true for eyes, which have independently evolved time and again. But the eukaryotic cell is a one-off innovation. Bacteria have repeatedly nudged along the path towards complexity. Some are very big (for microbes); others move in colonies that behave like single, many-celled creatures. But none of them have acquired the full suite of crucial features that define eukaryotes: large size, the nucleus, internal compartments, mitochondria, and more. As Nick Lane from University College London writes, “Bacteria have made a start up every avenue of eukaryotic complexity, but then stopped short.” Why? It is not for lack of opportunity. The world is swarming with countless prokaryotes that evolve at breathtaking rates. Even so, they were not quick about inventing eukaryotic cells. Fossils tell us that the oldest bacteria arose between 3 and 3.5 billion years ago, but there are no eukaryotes from before 2.1 billion years ago. Why did the prokaryotes remain as simple cells for so damn long? There are many possible explanations, but one of these has recently gained a lot of ground. It tells of a prokaryote that somehow found its way inside another, and formed a lasting partnership with its host. This inner cell—a bacterium—abandoned its free-living existence and eventually transformed into the mitochondria. These internal power plants provided the host cell with a bonanza of energy, allowing it to evolve in new directions that other prokaryotes could never reach. If this story is true, and there are still those who doubt it, then all eukaryotes—every flower and fungus, spider and sparrow, man and woman—descended from a sudden and breathtakingly improbable merger between two microbes. They were our great-great-great-great-...-great-grandparents, and by becoming one, they laid the groundwork for the life forms that seem to make our planet so special. The world as we see it (and the fact that we see it at all; eyes are a eukaryotic invention) was irrevocably changed by that fateful union—a union so unlikely that it very well might not have happened at all, leaving our world forever dominated by microbes, never to welcome sophisticated and amazing life like trees, mushrooms, caterpillars, and us.
IN 1905, the Russian biologist Konstantin Mereschkowski first suggested that some parts of eukaryotic cells were once endosymbionts—free-living microbes that took up permanent residence within other cells. He thought the nucleus originated in this way, as did the chloroplasts that allow plant cells to harness sunlight. He missed the mitochondria, but the American anatomist Ivan Wallin pegged them for endosymbionts in 1923. These ideas were ignored for decades until an American biologist—the late Lynn Margulis—revived them in 1967. In a radical paper, she made the case that mitochondria and chloroplasts were once free-living bacteria that had been sequentially ingested by another ancient microbe. That is why they still have their own tiny genomes and why they still superficially look like bacteria. Margulis argued that endosymbiosis was not a crazy, oddball concept—it was one of the most important leitmotivs in the eukaryotic opera. The paper was a tour de force of cell biology, biochemistry, geology, genetics, and paleontology. Its conclusion was also grossly unorthodox. At the time, most people believed that mitochondria had simply come from other parts of the cell.
“[Endosymbiosis] was taboo,” says Bill Martin from Heinrich Heine University Düsseldorf, in Germany. “You had to sneak into a closet to whisper to yourself about it before coming out again.” Margulis’ views drew fierce criticism, but she defended with equal vigor. Soon she had the weight of evidence behind her. Genetic studies, for example, showed that mitochondrial DNA is similar to that of free-living bacteria. Now, very few scientists doubt that mergers infused the cells of every animal and plant with the descendants of ancient bacteria. But the timing of that merger, the nature of its participants, and its relevance to the rise of eukaryotes are all still hotly debated. In recent decades,origin stories for the eukaryotes have sprouted up faster than old ones could be tested, but most fall into two broad camps. The first—let’s call it the “gradual-origin” group—claimed that prokaryotes evolved into eukaryotes by incrementally growing in size and picking up traits like a nucleus and the ability to swallow other cells. Along the way, these proto-eukaryotes gained mitochondria, because they would regularly engulf bacteria. This story is slow, steady, and classically Darwinian in nature. The acquisition of mitochondria was just another step in a long, gradual transition. This is what the late Margulis believed right till the end. The alternative—let’s call it the “sudden-origin” camp—is very different. It dispenses with slow, Darwinian progress and says that eukaryotes were born through the abrupt and dramatic union of two prokaryotes. One was a bacterium. The other was part of the other great lineage of prokaryotes: the archaea. (More about them later.) These two microbes look superficially alike, but they are as different in their biochemistry as PCs and Macs are in their operating systems. By merging, they created, in effect, the starting point for the first eukaryotes. Bill Martin and Miklós Müller put forward one of the earliest versions of this idea in 1998. They called it the hydrogen hypothesis. It involved an ancient archaeon that, like many modern members, drew energy by bonding hydrogen and carbon dioxide to make methane. It partnered with a bacterium that produced hydrogen and carbon dioxide, which the archaeon could then use. Over time, they became inseparable, and the bacterium became a mitochondrion. There are many variants of this hypothesis, which differ in the reasons for the merger and the exact identities of the archaeon and the bacterium that were involved. But they are all united by one critical feature setting them apart from the gradual-origin ideas: They all say that the host cell was still a bona fide prokaryote. It was an archaeon, through and through. It had not started to grow in size. It did not have a nucleus. It was not on the path to becoming a eukaryote; it set off down that path because it merged with a bacterium. As Martin puts it, “The inventions came later.” This distinction could not be more important. According to the sudden-origin ideas, mitochondria were not just one of many innovations for the early eukaryotes. “The acquisition of mitochondria was the origin of eukaryotes,” says Lane. “They were one and the same event.” If that is right, the rise of the eukaryotes was a fundamentally different sort of evolutionary transition than the gradual changes that led to the eye, or photosynthesis, or the move from sea to land. It was a fluke event of incredible improbability—one that, as far as we know, only happened after a billion years of life on Earth and has not been repeated in the 2 billion years since. “It’s a fun and thrilling possibility,” says Lane. “It may not be true, but it’s beautiful.” IN 1977, microbiologist Carl Woese had the bright idea of comparing different organisms by sequencing their genes. This is an everyday part of modern biology, but at the time, scientists relied on physical traits to deduce the evolutionary relationships between different species. Comparing genes was bold and new, and it would play a critical role in showing how complicated life like us—the eukaryotes—came to be. Woese focused on 16S rRNA, a gene that is involved in the essential task of making proteins and is found in all living things. Woese reasoned that as organisms diverge into new species, their versions of rRNA should become increasingly dissimilar. By comparing the gene across a range of prokaryotes and eukaryotes, the branches of the tree of life should reveal themselves. They did, but no one expected the results. Woese’s tree had three main branches. Bacteria and eukaryotes sat on two of them. But the third consisted of an obscure bunch of prokaryotes that had been found in hot, inhospitable environments. Woese called them archaea, from the Greek word for ancient. Everyone had taken them for obscure types of bacteria, but Woese’s tree announced them as a third domain of life. It was as if everyone was staring at a world map, and Woese had politely shown that a full third of it had been folded underneath. In Woese’s classic three-domain tree, the eukaryotes and archaea are sister groups. They both evolved from a shared ancestor that split off from the bacteria very early in the history of life on Earth. But this tidy picture started to unravel in the 1990s, as the era of modern genetics kicked into high gear and scientists started sequencing more eukaryotic genes. Some were indeed closely related to archaeal genes, but others turned out to be more closely related to bacterial ones. The eukaryotes turned out to be a confusing hodgepodge, and their evolutionary affinities kept on shifting with every new sequenced gene. In 2004, James Lake changed the rules of engagement. Rather than looking at any single gene, he and his colleague Maria Rivera compared the entire genomes of two eukaryotes, three bacteria, and three archaea. Their analysis supported the merger-first ideas: They concluded that the common ancestor of all life diverged into bacteria and archaea, which evolved independently until two of their members suddenly merged. This created the first eukaryotes and closed what now appeared to be a “ring of life.” Before that fateful encounter, life had just two major domains. Afterward, it had three. Rivera and Lake were later criticized for only looking at seven species, but no one could possibly accuse Irish evolutionary biologist James McInerney of the same fault. In 2007, he crafted a super-tree using more than 5,700 genes from across the genomes of 168 prokaryotes and 17 eukaryotes. His conclusion was the same: Eukaryotes are merger organisms, formed through an ancient symbiosis between a bacterium and an archaeon. The genes from these partners have not integrated seamlessly. They behave like immigrants in New York’s Asian and Latino communities, who share the same city but dominate different areas. For example, they mostly interact with their own kind: archaeal genes with other archaeal genes, and bacterial genes with bacterial genes. “You’ve got two groups in the playground and they’re playing with each other differently, because they’ve spent different amounts of time with each other,” says McInerney. They also do different jobs. The archaeal genes are more likely to be involved in copying and making use of DNA. The bacterial genes are more involved in breaking down food, making nutrients, and the other day-to-day aspects of being a microbe. And although the archaeal genes are outnumbered by their bacterial neighbors by 4 to 1, they seem to be more important. They are nearly twice as active. They produce proteins that play more central roles in their respective cells. They are more likely to kill their host if they are mistakenly deleted. Over the last four years, McInerney has found this same pattern again and again, in yeast, in humans, in dozens of other eukaryotes. This all makes sense if you believe the sudden-origin idea. When those ancient partners merged, the immigrant bacterial genes had to be integrated around a native archaeal network, which had already been evolving together for countless generations. They did integrate, and while many of the archaeal genes were displaced, an elite set could not be ousted. Despite 2 billion years of evolution, this core network remains, and retains a pivotal role out of all proportion to their small number. THE SUDDEN-ORIGIN hypothesis makes one critical prediction: All eukaryotes must have mitochondria. Any exceptions would be fatal, and in the 1980s, it started to look like there were exceptions aplenty. If you drink the wrong glass of water in the wrong part of the world, your intestines might become home to a gut parasite called Giardia. In the weeks that follow, you can look forward to intense stomach cramps and violent diarrhea. Agony aside, Giardia has a bizarre and interesting anatomy. Itconsists of a single cell that looks like a malevolent teardrop with four tail-like filaments. Inside, it has not one nucleus but two. It is clearly a eukaryote. But it has no mitochondria.
Mitochondria (left) are domesticated versions of bacteria (right) that now provide the cells of every animal, plant and fungus with energy. ShutterstockThere are at least a thousand other single-celled eukaryotes, mostly parasites, which also lack mitochondria. They were once called archezoans, and their missing power plants made them focal points for the debate around eukaryotic origins. They seemed to be living remnants of a time when prokaryotes had already turned into primitive eukaryotes, but before they picked up their mitochondria. Their very existence testified that mitochondria were a late acquisition in the rise of eukaryotes, and threatened to deal a knockout blow to the sudden-origin tales.
That blow was deflected in the 1990s, when scientists slowly realized that Giardia and its ilk have genes that are only ever found in the mitochondria of other eukaryotes. These archezoans must have once had mitochondria, which were later lost or transformed into other cellular compartments. They aren’t primitive eukaryotes from a time before the mitochondrial merger—they are advanced eukaryotes that have degenerated, just as tapeworms and other parasites often lose complex organs they no longer need after they adopt a parasitic way of life. “We’ve yet to find a single primitive, mitochondria-free eukaryote,” says McInerney, “and we’ve done a lot of looking.” With the archezoan club dismantled, the sudden-origin ideas returned to the fore with renewed vigor. “We predicted that all eukaryotes had a mitochondrion,” says Martin. “Everyone was laughing at the time, but it’s now textbook knowledge. I claim victory. Nobody’s giving it to me—except the textbooks.” IF MITOCHONDRIA were so important, why have they only evolved once? And for that matter, why have eukaryotes only evolved once? Nick Lane and Bill Martin answered both questions in 2010, in a bravura paper called, “The energetics of genome complexity,” published in Nature. In a string of simple calculations and elegant logic, they reasoned that prokaryotes have stayed simple because they cannot afford the gas-guzzling lifestyle that all eukaryotes lead. In the paraphrased words of Scotty: They cannae do it, captain, they just don’t have the power. Lane and Martin argued that for a cell to become more complex, it needs a bigger genome. Today, for example, the average eukaryotic genome is around 100–10,0001 times bigger than the average prokaryotic one. But big genomes don’t come for free. A cell needs energy to copy its DNA and to use the information encoded by its genes to make proteins. The latter, in particular, is the most expensive task that a cell performs, soaking up three-quarters of its total energy supply. If a bacterium or archaeon was to expand its genome by 10 times, it would need roughly 10 times more energy to fund the construction of its extra proteins. One solution might be to get bigger. The energy-producing reactions that drive prokaryotes take place across their membranes, so a bigger cell with a larger membrane would have a bigger energy supply. But bigger cells also need to make more proteins, so they would burn more energy than they gained. If a prokaryote scaled up to the same size and genome of a eukaryotic cell, it would end up with 230,000 times less energy to spend on each gene! Even if this woefully inefficient wretch could survive in isolation, it would be easily outcompeted by other prokaryotes. Prokaryotes are stuck in an energetic canyon that keeps them simple and small. They have no way of climbing out. If anything, evolution drives them in the opposite direction, mercilessly pruning their genomes into a ring of densely packed and overlapping genes. Only once did a prokaryote escape from the canyon, through a singular and improbable trick—it acquired mitochondria. Mitochondria have an inner membrane that folds in on itself like heavily ruched fabric. They offer their host cells a huge surface area for energy-producing chemical reactions. But these reactions are volatile, fickle things. They involve a chain of proteins in the mitochondrial membranes that release energy by stripping electrons from food molecules, passing them along to one another, and dumping them onto oxygen. This produces high electric voltages and unstable molecules. If anything goes wrong, the cell can easily die. But mitochondria also have a tiny stock of DNA that encodes about a dozen of the proteins that take part in these electron-transfer chains. They can quickly make more or less of any of the participating proteins, to keep the voltages across their membranes under check. They supply both power and the ability to control that power. And they do that without having to bother the nucleus. They are specialized to harness energy. Mitochondria are truly the powerhouse of the eukaryotic cell. “The command center is too bureaucratic and far away to do anything,” says Lane. “You need to have these small teams, which have limited powers but can use them at their discretion to respond to local situations. If they’re not there, everything dies.” Prokaryotes do not have powerhouses; they are powerhouses. They can fold their membranes inwards to gain extra space for producing energy, and many do. But they do not have the secondary DNA outposts that produce high-energy molecules so the central government (the nucleus) has the time and energy to undertake evolutionary experiments. The only way to do that is to merge with another cell. When one archaeon did so, it instantly leapt out of its energetic canyon, powered by its new bacterial partner. It could afford to expand its genome, to experiment with new types of genes and proteins, to get bigger, and to evolve down new and innovative routes. It could form a nucleus to contain its genetic material, and absorb other microbes to use as new tiny organs, like the chloroplasts that perform photosynthesis in plants. “You need a mitochondrial level of power to finance those evolutionary adventures,” says Martin. “They don’t come for free.” Lane and Martin’s argument is a huge boon for the sudden-origin hypothesis. To become complex, cells need the stable, distributed energy supply that only mitochondria can provide. Without these internal power stations, other prokaryotes, for all their evolutionary ingenuity, have always stayed as single, simple cells. The kind of merger that creates mitochondria seems to be a ludicrously unlikely event. Prokaryotes have only managed it once in more than 3 billion years, despite coming into contact with each other all the time. “There must have been thousands or millions of these cases over evolutionary time, but they’ve got to find a way of getting along, of reconciling and co-adapting to each other,” says Lane. “That seems to be genuinely difficult.” This improbability has implications for the search for alien life. On other worlds with the right chemical conditions, Lane believes that life would be sure to emerge. But without a fateful merger, it would be forever microbial. Perhaps this is the answer to the Fermi paradox—the puzzling contradiction between the high apparent odds that intelligent life would exist elsewhere among the billions of planets in the Milky Way, and our inability to find any signs of such intelligence. As Lane wrote in 2010, “The unavoidable conclusion is that the universe should be full of bacteria, but more complex life will be rare.” And if intelligent aliens did exist, they would probably have something like mitochondria, too. THE ORIGIN of eukaryotes is, by no means, a settled matter of fact. Ideas have waxed and waned in influence, and although many lines of evidence currently point to a sudden origin, there is still plenty of dissent. Some scientists support radical notions like the idea that prokaryotes are versions of eukaryotes that evolved to greater simplicity, rather than their ancestors. Others remain stalwart devotees of Woese’s tree. Writing in 2007, Anthony Poole and David Penny accused the sudden-origin camp of pushing “mechanisms founded in unfettered imagination.” They pointed out that archaea and bacteria do not engulf one another—that’s a hallmark of eukaryotes. It is easy to see how a primitive eukaryote might have gained mitochondria by engulfing a bacterium, but very hard to picture how a relatively simple archaeon did so. This powerful retort has lost some of its sting thanks to a white insect called the citrus mealybug. Its cells contain a bacterium called Tremblaya, and Tremblaya contains another bacterium called Moranella. Here is a prokaryote that somehow has another prokaryote living inside it, despite its apparent inability to engulf anything. Still, the details of how the initial archaeon-bacterium merger happened are still a mystery. How did one get inside the other? What sealed their partnership—was it hydrogen, as Martin and Müller suggested, or something else? How did they manage to stay conjoined? “I think we have the roadmap right, but we don’t have all the white lines and the signposts in place,” says Martin. “We have the big picture but not all the details.” Perhaps we will never know for sure. The origin of eukaryotes happened so far back in time that it’s a wonder we have even an inkling of what happened. Dissent is inevitable; uncertainty, guaranteed.
“You can’t convince everyone about anything in early evolution, because they hold to their own beliefs,” says Martin. “But I’m not worried about trying to convince anyone. I’ve solved these problems to my own satisfaction and it all looks pretty consistent. I’m happy.” ~ Ed Yong is an award-winning science writer. His work has appeared in Wired, Nature, the BBC, New Scientist, the Guardian, the Times, Aeon, Discover, Scientific American, The Scientist, the BMJ, Slate, and more. This article originally appeared in our “Mergers & Acquisitions” issue in February 2014.



No comments:
Post a Comment